
2) 팽윤에 의한 변형성능의 변화
목재는 물, 암모니아 혹은 저분자의 알콜, 페놀, 아민 등의 극성 기체나 액체를 흡착함으로써 팽윤(膨潤, swelling)한다. 이와 같은 팽윤제는 목재를 구성하고 있는 고분자의 분자 사슬 사이에 들어가 분자 간 결합을 약하게 하므로 외력이 가해질 때 분자사슬 상호 간의 어긋남에 의해 변형이 쉽게 된다. 이 상태에서 온도를 높이면 변형은 더욱 쉽게 일어난다. 즉, 목재는 팽윤에 의해서 탄성률이 작아지며 연화가 개시되는 온도도 낮아지게 진다. 그 정도는 팽윤제의 종류, 함유량(팽윤율)에 따라 다르다. 일반적으로 암모니아, 포름아마이드, 아민류에 의한 목재의 탄성률이나 연화가 개시되는 온도 저하는 물이나 알콜류에 의한 저하보다도 현저하다. 팽윤목재는 어떤 전이영역(轉移領域, transition region)을 사이에 두고 탄성률이 약 1/10로 저하하는데 불과하다.
이와 같은 변형성능에 미치는 팽윤효과 외에 일정한 외력이 가해진 상태에서 습윤목재(濕潤木材)가 건조되거나 온도가 상승할 때 목재의 함수율이나 온도가 일정할 때에 비하여 현저하게 변형성능이 향상되는 것으로 알려져 있다. 그 양은 초기 변형량의 3배 이상에 달한다. 게다가 변형량의 대부분은 외력을 제거한 후에도 영구변형으로서 남는다. 이와 같은 현상은 물 이외의 팽윤제로 팽윤시킨 목재에 대해서도 확인되고 있다.
3) 목재 구성성분의 가소화 효과
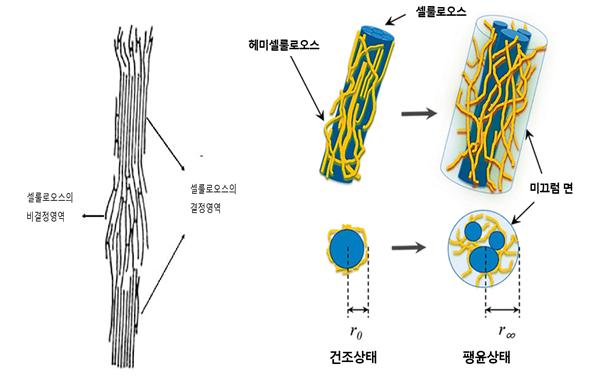
셀룰로오스(cellulose, 纖維素)의 비결정영역이나 헤미셀룰로오스(hemicellulose, 半纖維素 )는 물이나 기타 팽윤제에 대하여 강한 친화성을 가지므로 팽윤에 크게 기여하게 된다. 물분자가 셀룰로오스의 결정영역에 들어갈 수 없는 것으로 알려져 있는 데 반해 액체암모니아나 아민은 셀룰로오스의 결정영역에까지 들어가 미셀(micelle, 셀룰로오스의 정렬된 영역)내에서 팽윤을 일으키므로 외력이 가하여질 때 물에서 팽윤하는 이상으로 셀룰로오스 분자 상호의 미끄럼(어긋남, slip)이 용이해지는 것으로 생각된다. 그러나 셀룰로오스 미셀 내 팽윤은 목재 가소화의 한 인자이긴 하나 반드시 유리한 조건만은 아니다.
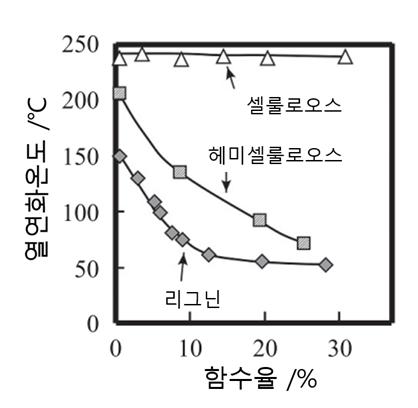
리그닌(lignin, 木質素)은 라틴어인 "lignum(목재 또는 나무)"이라는 용어에서 유래한 것으로 목재의 가소화와 관련하여 매우 중요한 성분이다. 최근 리그닌과 다당류의 상호작용에 대한 고체 NMR 분광법 연구에서는 리그닌이 스스로 응집하며(self-aggregate) 셀룰로오스와 밀접하게 결합하지 않은 자일란(xylan)에 대한 광범위한 표면 접촉을 가진 매우 소수성이고 동적으로 독특한 나노영역을 형성한다는 것을 시각적으로 잘 보여주고 있다(Xue Kang et al. 2019. Nature Communications 10: 347). 셀룰로오스의 연화점은 물분자 흡착의 유무와 관계없이 약 230℃인데 비하여 리그닌의 연화점은 수분의 존재에 의해 190℃에서 70~116℃로까지 낮아진다. 리그닌은 암모니아와도 강한 친화성을 가지고 있다. 리그닌의 용해성과 팽윤성은 용매의 수소결합 능력의 증대에 따라 커지나, 그 최대치는 응집에너지 밀도(cohesive energy density)가 10~12일 경우로 암모니아는 물에 비하여 이 최고치에 가깝다.
이상에서 목재를 팽윤제로 처리할 때의 소성변형은 셀룰로오스의 비결정영역, 헤미셀룰로오스와 리그닌의 분자 상호간 어긋남 또는 이들 성분의 세포벽 중에서의 분포로부터 이웃한 세포 사이의 세포간층(細胞間層, intercellular layer, 일명 중간층, 그림 7)에서의 세포 상호간의 어긋남이 중요한 인자로 작용한다.
리그노셀룰로오스 식물 세포벽은 주로 셀룰로오스, 헤미셀룰로오스 및 리그닌으로 구성된다. 셀룰로오스는 β-(1-4) 배열로 글리코사이드로 연결된 D-글루코스 단량체만으로 구성된 단일 다당류(homopolysaccharide)이다. 반복단위는 셀로비오스이다. 여러 선형 셀룰로오스 사슬은 수소결합으로 안정화된 기본 원섬유(elementary fibril)를 형성한다. 여러 묶음의 셀룰로오스 섬유가 응집하여 거대섬유(macrofibril)를 형성한다.
헤미셀룰로오스는 셀룰로오스와 달리 글루코스와 자일로스, 만노스 및 아라비노스와 같은 식물 종에 따라 다른 오탄당(pentose)으로 구성된 곁가지를 가지는 분지형(分枝形, branched) 헤테로다당류이다. 리그닌의 단위체 구성 요소는 탄소-탄소 및 에테르 결합으로 연결된 코니페릴알코올, ρ-쿠마릴알코올 및 시나필알코올이다. 세 가지 중합체인 셀룰로오스, 헤미셀룰로오스 및 리그닌은 난분해성이 매우 높은 복합구조인 리그노셀룰로오스를 형성한다.
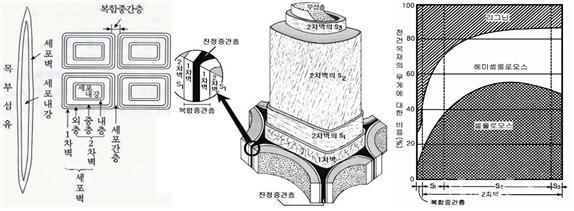
식물에서 셀룰로오스는 로제트 말단 복합체(rosette terminal complexes, RTCs)에 의해 세포막에서 생합성된다. 로제트 말단 복합체는 세포막 안쪽 면의 미세공극을 통해 셀룰로오스 사슬을 바깥으로 뻗어나간다. 헤미세룰로오스는 셀룰로오스 다발을 감싼다. 셀룰로오스와 헤미셀룰로오스의 다당류 다발 사이에 리그닌이 헤미셀룰로오스와 접착제처럼 결합하여 세포벽을 완성하고 세포벽의 두께를 확대해 나간다. 세포벽은 중간층(주로 펙틴) → 1차 세포벽 → 2차 세포벽(목질화)의 순서로 형성된다. 세포와 세포 사이의 중간층과 1차벽에는 주로 펙틴(pectin)이 다량 존재하고, 2차벽에는 리그닌이 생겨 셀롤로오스와 헤미셀룰로오스를 접착하여 세포벽을 단단하게 한다. 2차 세포벽은 일반적으로 외부의 벽면 1층(S1), 중앙의 벽면 2층(S2), 가장 안쪽의 벽면 3층(S3), 이렇게 세 개의 층이 있다. 각 층엔 셀룰로스 미소섬유(microfibril)가 서로 다른 방향으로 구성되어 있다. S2층은 세포벽의 75%까지 차지할 수 있다. 중간층과 1차 세포벽은 단지 인접한 세포를 분리하는 얇은 막을 형성한다. 중간층과 1차벽을 합친 진정중간층에는 리그닌 성분이 대부분을 차지하고 있다. 2차벽에도 리그닌이 20% 정도 포함되어 있다(그림 7, 8).
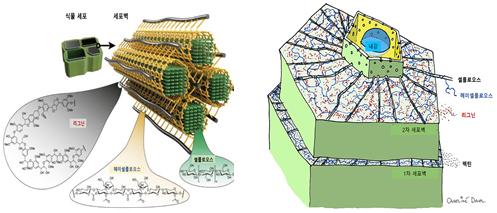
따라서 목재의 유동성을 확보하기 위해서는 2차벽의 50%를 차지하는 셀룰로오스를 에스테르화 등에 의해 단단히 뭉쳐있는 결정구조를 용융구조로 바꾸어야 한다. 그리고 헤미셀룰로오스와 리그닌은 흡수와 가열에 의해 유동성을 향상시킬 수 있다. 목재의 가소화에 있어서 중간층의 리그닌이 흡수·가열에 의해 용융되면서 세포층 간 미끄러짐이 먼저 발생하고, 이어 세포벽 내의 상기 3성분(셀룰로오스, 헤미세룰로오스 및 리그닌)이 첨가제 반응(에스테르화, 에테르화 등)에 의해 열가소화되면서 목재 전체가 완전히 유동성을 발현하는 것으로 생각된다.